Research
Research
私たちは、生き物がその個体性をどのように獲得し、維持していくのかということについて、環形動物ゴカイや棘皮動物ヒトデの生殖・免疫・形態形成といった現象に着目して研究を展開しています。個体や細胞レベルでの行動や形態観察だけでなく、得られた観察結果がどのようなメカニズムに裏打ちされているのかまで明らかにするために、トランスクリプトーム解析や分子機能の解析も精力的に進めています。
主要なテーマ
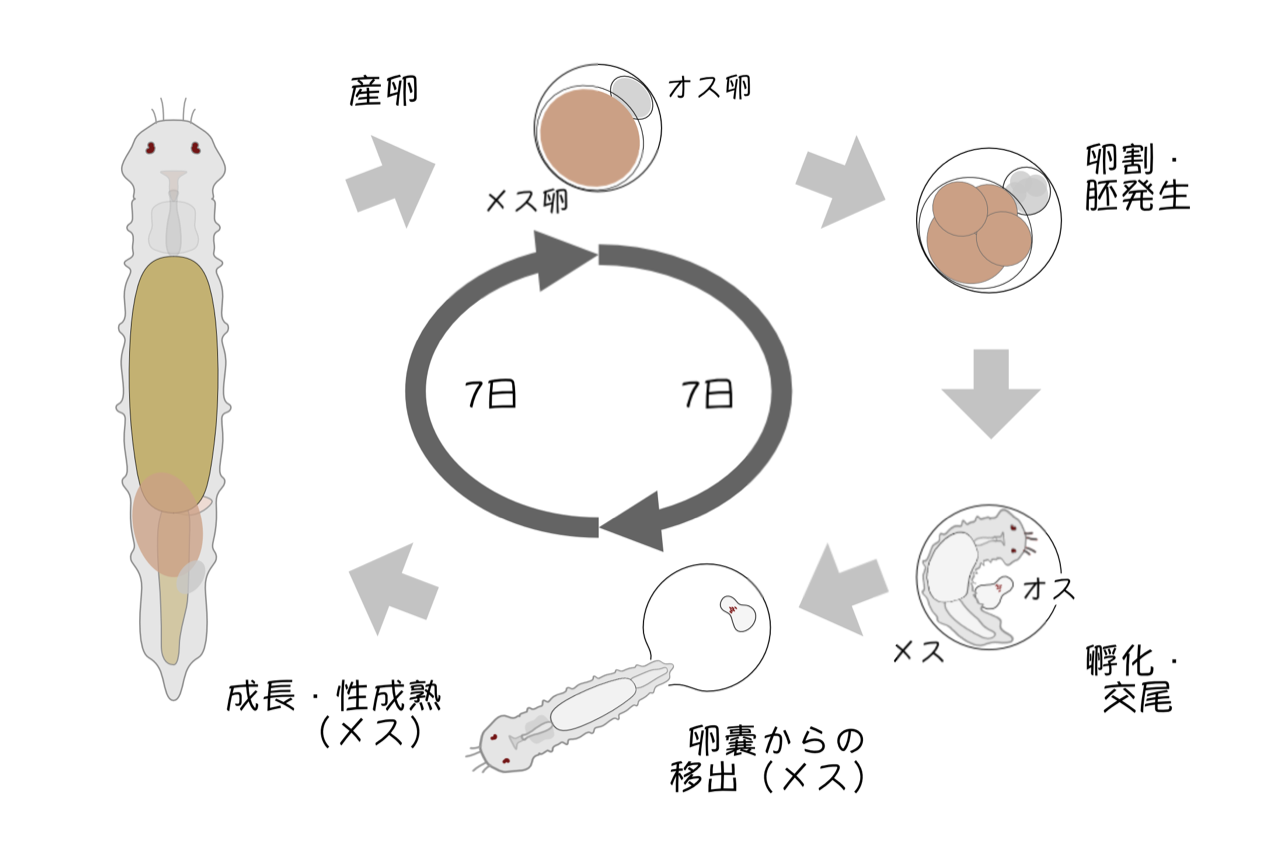
<クリックで拡大>
Dinophilus 属のゴカイは短いライフサイクル(20℃で約2週間)、幼形成熟、卵嚢内での同胞生殖、極端な性的2形(受精前から雌雄で卵サイズが異なり、矮性のオスは卵嚢内で交尾して一生を終える)といった生活史上の特徴を持っています(図1)。飼育下で容易に継代飼育でき、性質の揃った胚や個体を年中得られることから、古くから環境毒性試験の材料などに用いられてきました。しかし、その生殖に関しては実はよくわかっていません。メスの体内で卵形成・卵成熟のどの時点で精子の侵入が起きているのか、多精受精や減数分裂はメスの体内でどのようにして抑制されているのか、産卵に伴う卵の活性化(減数分裂とそれに続く卵割の開始)はどのように制御されているのか、オスの発生はどのような修飾をうけているのか、など様々な問題が残されています。また、予備的な研究からメスは孵化して性的に成熟した後も交尾能力を維持していることがわかってきました。複数回交尾が可能なのであれば、どのオスの精子が受精に使用されるのかという点も興味深い問題です。生き物のハンドリング自体未だ手探りの段階ですが、これらの問題についてアプローチする手立てを検討しています。
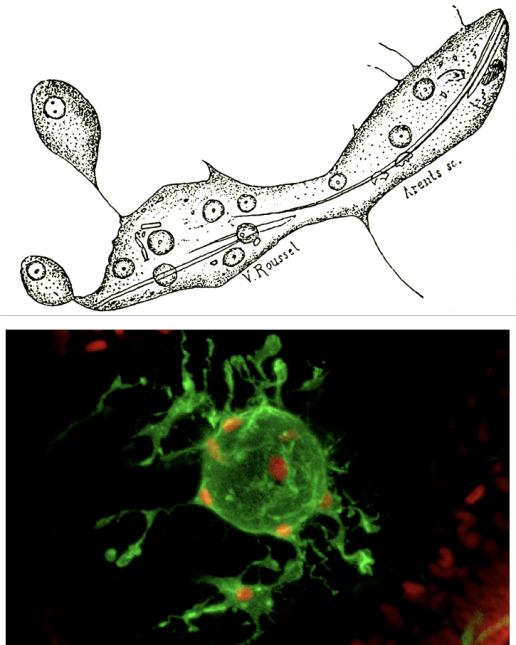
<クリックで拡大>
ヒトデが属する棘皮動物は、わたしたちを含む脊椎動物に繋がる後口動物の基部に位置しており、系統進化を考える上で非常に重要な動物です。ヒトデの幼生の免疫システムは、間充織細胞と呼ばれる1種類の免疫細胞で構築されています。実はこの間充織細胞は、免疫を担う細胞が存在するということを私たち人類に教えてくれた、記念碑的な細胞です(Metchnikoff, 1891; 図2)。間充織細胞は活発な貪食能力を持っており、外界から侵入した異物や、体内で死んだ細胞などを貪食し、除去します。このような貪食細胞は全ての多細胞動物に存在しているため、原始的な動物における貪食細胞の機能と、それを支えるメカニズムの解析を通して、免疫システムがどのように進化してきたかという問題を考えるヒントが得られると期待されます。
ヒトデの幼生には免疫細胞が1種類しかいないため、その免疫システムは多細胞動物において最もシンプルなシステムだと考えられます。わたしたちは、それがどのようなシステムかを明らかにするために、間充織細胞がどのように情報を伝達し、それがどのように細胞内に伝えられ、どのような免疫行動を制御しているのかという一連の流れについて研究を進めています。一方、間充織細胞の免疫機能を支える分子の一部は、胚や幼生の発生、形態形成にも関与します。個体が、このような分子をどのように使い分けているのかについても興味を持って研究しています。
間充織細胞の免疫行動制御メカニズムに関する私たちの報告は、以下のサイトでも紹介されています。→ ![]()
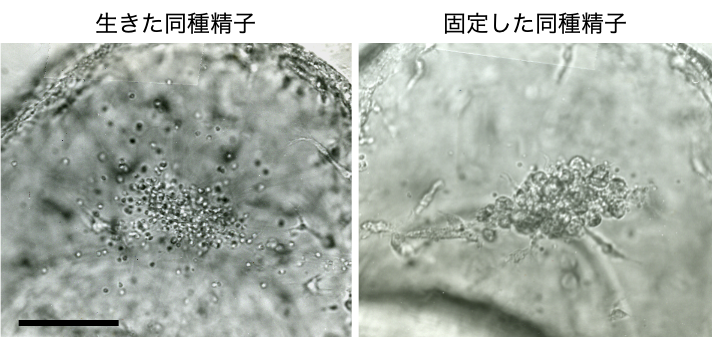
<クリックで拡大>
免疫システムとは、自己と非自己を識別し、非自己を体内から排除することで個体の自己同一性を維持するシステムです。「非自己」とは「自分ではないもの」であるため、非自己を識別するためには「自己」という存在が確立されている必要があります。つまり、免疫システムの進化を考えることは、「自己とは何か?」という問題を考えることでもあります。
ヒトデとして一般的にイメージされる星型の成体(大人)ヒトデでは、わたしたち同様に移植拒絶反応が存在することが古くから知られています。これは、ヒトデの成体の免疫システムは、同種異個体を非自己と認識していることを意味します。一方で、わたしたちは、成体になる前の幼生期のヒトデは同種異個体を拒絶せず、生きた同種細胞であれば免疫応答を生じないということを発見しました(Furukawa et al., 2009)。実際、幼生期に存在しない同種の精子を体内に注射しても、精子が生きている間は間充織細胞に貪食されることはありません(図3)。
無脊椎動物は、わたしたちが持っている抗体のような、特定の異物に対して特異的に反応することができるツールを持っていません。したがって、無脊椎動物の免疫システムは、「自己」を特異的に認識することで、認識できないものを「非自己」とみなしていると考えられています。この認識様式を「missing-self(自己喪失)認識」と言います。ヒトデの成体における同種異個体からの移植拒絶反応も、「missing-self認識」による免疫応答の結果だと言えるでしょう。一方、この考えをヒトデの幼生の免疫システムに当てはめると、幼生の間充織細胞による認識様式は、「同種」を特異的に認識し、認識できないものを「非自己」とみなす「missing-species(種喪失)認識」だと解釈することができます。わたしたちは、この「自己」=「同種」というシステムを裏打ちする分子の同定を目指すとともに、幼生と成体の免疫システムの認識対象の差にも興味を持って研究を進めています。
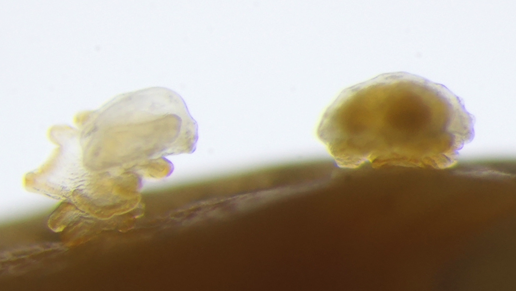
<クリックで拡大>
幼生と成体の自己認識様式の違いは、ヒトデにおいて個体性がいつどのように確立されるのかという問題に繋がります。わたしたちは、ヒトデの変態現象にこの問題を解き明かすヒントがあるのではないかと考えています。そこで、変態の各ステージでトランスクリプトームデータを取得し、免疫関連分子の発現量変化に着目して解析を進めています。
同時に、このヒトデの変態現象には別の視点からも着目しています。ヒトデの幼生はブラキオラリア期になると、変態する能力を獲得します。しかし、ブラキオラリア幼生になるだけでは変態することはできず、変態後の生息に適した環境が必要です。生息に適した環境に到達すると頭部の付着器官を介して岩場などに固着し、そこでようやく変態を開始します(図4)。この変態開始の合図となっているのは、固着する基質表面に存在するバクテリアからの刺激です。個体性を確立する上で重要なイベントであると推測される変態が、外界のバクテリア、つまり「非自己」による刺激で生じるということに強く興味を持っています。
最近、別のグループの研究(Yamakawa et al., 2018)により、レチノイン酸の投与で、バクテリアが存在しなくてもヒトデが変態を開始できることが明らかになりました。この事実は、ヒトデが変態を開始するとき、バクテリア刺激の下流でレチノイン酸シグナルが働いていることを示唆しています。レチノイン酸には様々な生理活性があることが知られていますが、免疫システムにおいても重要な役割を担っていると考えられています。非自己に由来する変態誘起シグナルがヒトデの免疫システムにどのような影響を及ぼすのかという視点は、ヒトデの個体性確立システムを理解する上で有用な知見を提供してくれるだろうと期待しています。

<クリックで拡大>
1902年、Wilsonという人が、バラバラに解離したカイメンの細胞が、再び凝集して元の個体を再構築することを発見しました。その後、この再構築現象はイモリなどの脊椎動物でも観察され、多くの研究者の興味を惹いてきました。
再構築現象に対するわたしたちの大きな興味は、自然界では起こり得ない状況で、細胞の運命や形態形成がどのように制御されているのかという点です(図5)。わたしたちは、ヒトデの胚や幼生を用いて、いろいろなオリジナルの実験系を開発しながらこの問題に取り組んでいます。最近、再構築過程のトランスクリプトームデータを用いたネットワーク解析から、再構築の成立に必須の遺伝子を発見することができました(未発表)。この遺伝子を中心に、再構築過程の個体の各領域でどのような遺伝子ネットワークが存在するかを調べることで、再構築過程における個々の細胞のキャラクターの違いや、形態形成がどのように制御されているのかなどを追求していくことができるだろうと期待しています。
一方、全く別の視点で、再構築現象をツールとして用いる研究も進行中です。「ヒトデにおける自己・非自己認識」のセクションで述べたとおり、幼生の免疫システムは同種異個体を識別できませんが、成体になると識別可能になります。では、遺伝的に異なる同種の幼生の解離細胞を混ぜ合わせて再構築させた場合、その個体は変態して成体になることが出来るのでしょうか? 非常に興味深い問題ですが、これは近い将来明らかにすることができると思っています。
ヒトデの再構築現象に関する私たちの報告は、以下のサイトでも紹介されています。→ ![]()